la raíz
es un órgano generalmente subterráneo y carente de hojas que crece en dirección inversa al tallo y cuyas funciones principales son la fijación de la planta al suelo y la absorción de agua y sales minerales. La raíz está presente en todas las plantas vasculares exceptuando algunas pteridófitas que presentan rizoides y algunas plantas acuáticas.1 La raíz del embrión —llamada radícula— es la primera de las partes de la semilla que crece durante la germinación. La radícula, entonces, se desarrolla originando la raíz primaria con su tejido de protección en el ápice, denominada caliptra.
La radícula crece y se fija al suelo desde los primeros estadios del
crecimiento de la planta, con lo cual se garantiza el posterior
desarrollo de la misma. En las plantas monocotiledóneas, la radícula aborta en los estados iniciales del desarrollo, por lo que el sistema radical está conformado por raíces que surgen de la base del tallo, las que —por ese motivo— se denominan raíces adventicias.2 En las gimnospermas y dicotiledóneas
la raíz primaria produce, por alargamiento y ramificación, el sistema
radical alorrizo, caracterizado porque hay una raíz central, principal,
nítida y dominante sobre las raíces laterales. En las monocotiledóneas
y en las pteridófitas, en cambio, el sistema radical de la planta
adulta se forma por encima del lugar de origen de la raíz primaria que
aborta tempranamente. El sistema radical de estas plantas se denomina
homorrizo, fasciculado, en cabellera o fibroso, y está formado por un
conjunto de raíces adventicias y se halla profusamente ramificado.3Las raíces pueden experimentar modificaciones estructurales pronunciadas, que pueden ser consideradas, en la mayoría de los casos, como adaptaciones al medio ambiente, o bien, la consecuencia de una especialización funcional diferente a la función típica de este órgano. Entre éstas se encuentran las raíces reservantes y las raíces especializadas como órganos de sostén y fijación.4 5
La porción de suelo que envuelve a las raíces de las plantas se denomina rizosfera y es una zona donde se producen una serie de relaciones físicas y químicas que afectan a la estructura del suelo y a los organismos que viven en él, proporcionándole unas propiedades diferentes. La rizosfera normalmente ocupa entre unos cuantos milímetros o algunos centímetros alrededor de la raíz. Esta región se caracteriza por el aumento de la biomasa microbiana y de su actividad. La comunidad de la rizosfera consiste en una microbiota (bacterias, hongos y algas) y una micro y mesofauna (protozoos, nematodos, insectos y ácaros).6 Las micorrizas constituyen una simbiosis especialmente importante, que ocurre en la mayoría de los grupos de plantas vasculares. El término define a la simbiosis entre un hongo y las raíces de una planta. Como en otras relaciones simbióticas, ambos participantes obtienen beneficios. En este caso la planta recibe del hongo principalmente nutrientes minerales y agua y el hongo obtiene de la planta hidratos de carbono y vitaminas que él por sí mismo es incapaz de sintetizar mientras que ella lo puede hacer gracias a la fotosíntesis y otras reacciones internas.7 8 Los nódulos radicales son asociaciones simbióticas entre bacterias y plantas superiores. La más conocida es la de Rhizobium con especies de la familia de las leguminosas. La planta proporciona a la bacteria compuestos carbonados como fuente de energía y un entorno protector, y recibe nitrógeno en una forma utilizable para la formación de proteínas.9
Las raíces evolucionaron en los esporofitos de por lo menos dos linajes diferentes de las plantas vasculares durante su principal radiación adaptativa sobre la Tierra en el período Devónico inferior (hace unos 410 a 395 millones de años). Ese hecho ocurrió aproximadamente unos 15 millones de años después de la aparición de las traqueófitas y unos 50 millones de años después de las primeras embriófitas con afinidad presunta con las briófitas. Ambos grupos se conocen solo por sus esporas pero se supone que tenían algún órgano de anclaje al sustrato.10 Para el Devónico intermedio a tardío, la mayoría de los grupos de plantas desarrollaron un sistema radicular de alguna naturaleza.11 A medida que las raíces se hacían más largas, podían sustentar estructuras aéreas más altas y podían explorar el sustrato a mayor profundidad.12 Esta exploración más eficiente del suelo tuvo profundos efectos ecológicos: no solo permitió a las plantas la conquista de nuevos hábitats sino también la posterior colonización de los mismos por los animales y los hongos.13
Índice
Concepto
La raíz es el órgano generalmente subterráneo del cuerpo de las cormófitas, que se caracteriza por su crecimiento indefinido, su geotropismo positivo, su simetría en general radiada, la ausencia de yemas, hojas, nudos y entrenudos y por su especialización como órgano de anclaje, absorción de agua y sales minerales disueltas; de acumulación de diversas sustancias orgánicas y en ocasiones excepcionales como unidad de propagación.2 14Origen y distribución
La raíz se origina a partir de la radícula del embrión, o polo radical del eje embrionario, y se conoce como «raíz principal» o «raíz primaria». Es la primera de las partes del embrión que se desarrolla durante la germinación de la semilla. La radícula, entonces, con una cubierta en su punta llamada coleorriza, se desarrolla originando la raíz primaria con su tejido de protección en el ápice, denominada caliptra, cofia o pilorriza. La radícula crece y se fija al suelo desde los primeros estadios, de esta modo se garantiza el posterior crecimiento y desarrollo de la planta. En las plantas monocotiledóneas, la radícula aborta en estados tempranos de desarrollo, por lo que el sistema radical está conformado por raíces que surgen de la base del tallo, las que —por ese motivo— se denominan raíces adventicias.2 15La raíz está presente en todas las plantas vasculares exceptuando algunas pteridófitas —como las psilotáceas— que presentan rizoides. Ciertas espermatófitas especializadas carecen de raíz porque se atrofia el polo radical y el embrión no presenta radícula. Entre estos casos particulares se hallan varias especies de plantas acuáticas, tales como Wolffia, Utricularia y Ceratophyllum demersum, y de plantas epífitas como Tillandsia usneoides y algunas orquídeas. Algunas de estas especies pueden formar raíces adventicias para sustituir a la raíz principal.16 En Salvinia, una pteridófita acuática, la función de la raíz es desempeñada por hojas modificadas.3
Sistema radicular
El conjunto de raíces de una misma planta se denomina sistema radical o sistema radicular. Según su origen y desarrollo se distinguen dos tipos de sistemas radiculares, los cuales están asociados a grupos diferentes de plantas. En las gimnospermas y dicotiledóneas la raíz primaria produce, por alargamiento y ramificación, el sistema radical alorrizo, caracterizado porque hay una raíz central, principal, nítida y dominante sobre las raíces laterales, las que no son morfológicamente equivalentes. El sistema radical generalmente es unitario, presenta ramificación racemosa y acrópeta, es decir, que progresa hacia el ápice. En este sistema la raíz se dice axonomorfa o pivotante, tiene raíces de segundo a quinto orden y crecimiento secundario.1En las monocotiledóneas y en las pteridófitas, la raíz embrionaria por lo general muere pronto y el sistema radical de la planta adulta se forma por encima del lugar de origen de la raíz primaria. El sistema radical se denomina homorrizo, fasciculado, en cabellera o fibroso, y está formado por un conjunto de raíces adventicias y se halla profusamente ramificado.3 1
Funciones
La raíz cumple varias funciones en la planta. Por un lado, permite el anclaje o fijación de la planta al suelo. El tamaño relativo de las raíces determinan también la posibilidad de que una planta pueda tener un mayor o menor desarrollo del vástago aéreo. La raíz también permite la absorción del agua y de los nutrientes minerales disueltos en ella desde el suelo y su transporte al resto de la planta. Asimismo, la raíz es el soporte de asociaciones simbióticas complejas con varios tipos de microorganismos, tales como bacterias y hongos, que ayudan a la disolución del fósforo inorgánico del suelo, a la fijación del nitrógeno atmosférico y al desarrollo de las raíces secundarias.3 17 18Además de estas tres funciones que son generales para todas las plantas superiores, la raíz de algunas especies están especializadas en la acumulación o almacenamiento de reservas. Así, las plantas bienales como la zanahoria (Daucus carota) almacenan en la raíz durante el primer año reservas que utilizarán el segundo año para producir flores, frutos y semillas. En algunas plantas como Isoetes (una pteridófita) y Littorella (una dicotiledónea de la familia de las plantagináceas) las raíces transportan dióxido de carbono para la fotosíntesis, ya que sus hojas usualmente carecen de estomas.3
La raíz, por otro lado, tiene un papel fundamental en la creación y protección del suelo. Las moléculas y enzimas segregadas por las raíces y sus relaciones simbióticas contribuyen a la formación de suelo. Las raíces de numerosos árboles segregan ácidos orgánicos bastante potentes para disolver piedras calizas y liberar el calcio y otros minerales útiles. Además, las raíces de numerosas especies contribuyen a la fijación del suelo y a protegerlo de la erosión hídrica y eólica.18 5
Morfología

La proliferación del tejido meristemático o de crecimiento origina las células que, tras su diferenciación, forman los tejidos adultos de la raíz. Entre tales tejidos se encuentran el parénquima, los tejidos vasculares y, en aquellas raíces que se deben engrosar en años sucesivos, meristemas remanentes —como el cámbium y el felógeno— responsables del crecimiento secundario o crecimiento en grosor de la raíz.17
La forma general de las raíces es filamentosa, aún cuando también se puede presentar en forma cónica, oblonga, napiforme, tortuosa o laminar. Cada especie tiene una forma característica, la cual puede modificarse según las diferentes condiciones ambientales en las que la planta crezca. De hecho, su apariencia depende del tipo de sustrato en el que se desarrolle.
Ramificación de la raíz
Cada raíz tiene un número definido de filas de raíces laterales, los que se denominan «rizósticos». Las raíces con más de dos polos de xilema, forman tantas filas de raíces como polos hay. En varias familias de monocotiledóneas con raíces poliarcas —tales como las gramíneas, las juncáceas y las ciperáceas— las raíces laterales se forman frente a los polos de floema, mientras en las dicotiledóneas se forman frente a los polos de xilema. Las bromeliáceas constituyen una excepción dentro de las monocotiledóneas ya que sus raíces laterales se forman frente a los polos de xilema. Por otro lado, en las raíces diarcas, las raíces laterales se originan entre los polos de xilema y floema, razón por la cual presentan cuatro filas de raíces laterales, es decir el doble que el número de polos.19 5
Raíces adventicias
Las raíces adventicias son aquellas que no provienen de la radícula del embrión, sino que se originan en cualquier otro lugar de la planta, como por ejemplo en alguna porción del vástago, en tallos subterráneos y en raíces viejas. Pueden tener o no ramificaciones, pero tienen una forma y un tamaño relativamente homogéneo, formando sistemas radicales fibrosos. Generalmente no presentan crecimiento secundario. Su duración varía, en algunos pastos perennes pueden durar varios años.20 En muchas monocotiledóneas como la gramilla (Cynodon dactylon) y dicotiledóneas como la frutilla (Fragaria), que presentan tallos postrados, en cada nudo nace un fascículo de raíces adventicias.21
Algunas plantas con un solo tallo, como las palmeras Socratea y Pandanus, logran mayor estabilidad desarrollando raíces adventicias llamadas raíces fúlcreas o raíces zancos. Dichas raíces también se encuentran en gramíneas como el maíz y el sorgo. Son gruesas, se originan en los nudos basales y penetran al suelo donde cumplen una doble función: sostén y absorción.5

La propiedad de muchas especies de formar raíces adventicias a partir de los extremos cortados de sus tallos se utiliza como medio de multiplicación asexual por medio de esquejes. Algunas especies, tales como los sauces y los geranios, arraigan con mucha facilidad, mientras que otras, como las coníferas, casi nunca emiten raíces si los esquejes no son previamente sometidos a un tratamiento especial. Tal tratamiento de los esquejes involucra la aplicación de fitohormonas, compuestos que las plantas sintetizan de forma natural para estimular la formación de raíces nuevas. Casi todos los preparados comerciales de este tipo contienen ácido indolacético, uno de los estimulantes más conocidos para la formación de raíces. En ocasiones, las raíces pueden originarse de las hojas, como ocurre en la violeta africana, especie que se puede propagar sumergiendo en agua el borde cortado de una hoja. Las raíces de algunas plantas también emiten brotes; así, los tallos que se forman a distancias variables de la base del chopo negro brotan desde las raíces del árbol.22
Anatomía
Caliptra

La caliptra está formada por células parenquimáticas vivas que a menudo contienen almidón. Las células se disponen en hileras radiales, las células centrales forman un eje llamado columela. Las células apicales se diferencian en células periféricas que junto con las células epidérmicas secretan el «mucigel», sustancia viscosa compuesta principalmente por polisacáridos elaborados en los dictiosomas. El extremo de la raíz está revestido de mucigel, envoltura viscosa constituida por mucílago que la protege contra productos dañinos, previene la desecación, es la interfase de contacto con las partículas del suelo y proporciona un ambiente favorable a los microorganismos. Las células periféricas se desprenden a medida que la raíz se abre paso en el suelo.23
La caliptra provee de protección mecánica a las células meristemáticas cuando la raíz crece a través del suelo. Estas células son destruidas por el crecimiento de la raíz y la fricción con el suelo, pero son rápidamente reemplazadas por células nuevas generadas por división celular en la cara externa del meristema de la raíz. A pesar de que continuamente se forman nuevas células en la parte profunda de la caliptra, ésta no aumenta de tamaño porque las células externas se desprenden, se descaman, por gelificación de las laminillas medias. La caliptra también está implicada en la producción de mucílago, que es una substancia gelatinosa que cubre a las células meristemáticas recién formadas. Estas células contienen estatolitos, que son granos de almidón que se hallan dentro de la célula y son muy densos, por lo que se mueven en respuesta a la fuerza de la gravedad, proporcionando a la raíz la información necesaria para su crecimiento.23 17
Rizodermis

En la región adyacente a la caliptra las células de la rizodermis son pequeñas y con citoplasma denso, sin vacuolas. En raíces que conservan su epidermis por largo tiempo, reemplazándola tardíamente por peridermis, las paredes celulares pueden engrosarse, es decir pueden suberificarse (deponer suberina) o lignificarse (deponer lignina). Los pelos radicales se encuentran en la denominada «zona pilífera» de la raíz y son parte de la rizodermis. Pueden originarse en todas las células epidérmicas, en ciertas células llamadas tricoblastos, o en la capa subepidérmica. Son tubulosos, raramente ramificados, con una vacuola central gigantesca, con citoplasma parietal, el núcleo es poliploide y se dispone en el extremo celular que se va alargando. En general viven pocos días, si bien en las gramíneas, por ejemplo, los pelos absorbentes son a menudo persistentes; en Nardus stricta y en las especies de la familia de las compuestas pueden durar 3 a 4 años. La función de los pelos radiculares es la de aumentar la superficie de absorción de la raíz para la toma de agua y nutrientes en solución. Los pelos radicales no se desarrollan en las raíces de plantas hidrófitas, las cuales pueden absorber agua en toda su superficie. Cuando la raíz crece produce nuevos pelos radiculares para reemplazar a los que van muriendo.25 24 17
Muchas orquídeas (en particular, las orquídeas epífitas) y algunas otras monocotiledóneas, como las aráceas, ciperáceas y veloziáceas presentan una rizodermis especializada denominada velamen. El velamen consta de células muertas a la madurez con engrosamientos de lignina en la pared celular. El velamen constituye una vaina esponjosa y blanquecina que rodea por completo a la raíz. Si el tiempo está seco, sus células están llenas de aire; pero cuando llueve se llenan de agua. Según algunos autores el velamen es un tejido que absorbe agua, según otros nunca se ha observado el paso de agua del velamen al córtex de la raíz. Su función principal parece ser la de protección mecánica, además de impedir la excesiva pérdida de agua de la raíz en períodos de deficiencia hídrica.24 17
Córtex
El córtex es la región comprendida entre la rizodermis y el cilindro central y su función principal es la de almacenar sustancias de reserva, tales como el almidón. Las capas más externas del córtex, debajo de la epidermis, pueden diferenciarse como un tejido especializado, llamado exodermis. La capa más interna del córtex forma, a su vez, otra estructura especializada en las espermatófitas: la endodermis.26El córtex propiamente dicho (la zona comprendida entre exodermis y endodermis) tiene en general una estructura homogénea, si bien en algunas especies puede estar formado por varios tipos de células. Las raíces normalmente no presentan clorofila en el córtex, pero frecuentemente las células contienen almidón; pueden encontrarse idioblastos diversos, como por ejemplo células taníferas o cristalíferas; puede presentar estructuras secretoras como espacios intercelulares lisígenos o esquizógenos. En las raíces con crecimiento secundario de las gimnospermas y de las dicotiledóneas que desprenden pronto su córtex, éste es parenquimático. En las monocotiledóneas, en cambio, en las que el córtex se conserva durante mucho tiempo, el esclerénquima se presenta en abundancia. Este tejido puede tener disposición cilíndrica dentro de la exodermis o junto a la endodermis. También puede encontrarse colénquima. El córtex en las plantas acuáticas y palustres, como así también en las gramíneas de hábitats relativamente secos, está constituido por aerénquima. Las raíces aéreas de muchas familias de plantas epífitas, tales como las orquidáceas y las aráceas, presentan cloroplastos en las células periféricas del córtex.26 5
- Exodermis
Endodermis

Cuando se produce el crecimiento secundario, con la formación profunda de peridermis, la endodermis se separa de la raíz con el córtex. Cuando la peridermis se forma superficialmente, la endodermis se estira y se aplasta o se acomoda a la expansión del cilindro vascular por divisiones celulares anticlinales.27
Cilindro vascular

Con respecto a los tejidos vasculares, el floema forma cordones por debajo del periciclo. En las angiospermas está formado por tubos cribosos y células acompañantes; mientras que en las gimnospermas está formado por células cribosas y raramente se presentan fibras. El xilema está dispuesto también en cordones, los cuales alternan con los de floema y puede estar constituido por vasos y traqueidas. En ocasiones, el xilema no ocupa el centro del cilindro vascular; cuando lo hace, los elementos de mayor diámetro están en el centro. En ese caso, el cilindro vascular se describe como una actinostela como sucede por ejemplo en Clintonia, o protostela. Según el número de "polos" (cordones) de protoxilema se reconocen diferentes tipos de raíz. Con presentan solamente un polo, la raíz se denomina monarca (el caso de Trapa natans, una planta acuática); con dos, diarca (en las pteridófitas y dicotiledóneas tales como Daucus, Solanum y Linum); con tres, triarca (Pisum); con cuatro, tetrarca (Vicia, Ranunculus); con cinco, pentarca, y poliarca con varios polos. Las raíces de las gimnospermas son diarcas o poliarcas.27
Células del borde
Las células del borde son células vivas que se separan de la periferia de la raíz al sustrato. Inicialmente se llamaban "sloughed root cap" (caliptra desprendida) pero por definición ya no son parte de la caliptra así que se propuso el término de "células del borde"29 . La propuesta de que las células del borde y la caliptra son tipos celulares distintos está apoyada por la observación de que su perfil protéico es diferente30 . No todas las raíces presentan células del borde. En raíces con meristemo apical de tipo cerrado, su caliptra se descama por capas de células muertas mientras que las raíces con meristemo apical de tipo abierto si presentan células del borde31 . El tipo de meristemo apical de la raíz está definido por la forma en que se organizan las filas de células.En la punta de la raíz se encuentra una capa de células que se conoce como meristemo de la caliptra y entre sus funciones principales destaca la producción de células del borde. Después de que estas células se producen en el meristemo, diferencian progresivamente hasta llegar a la periferia de la raíz donde se desprendende hacia la rizosfera inmersas en mucílago29 . La producción de las células del borde está regulada por un inhibidor que ellas mismas producen. De esta manera su producción es un proceso autolimitante ya que entre más células del borde se acumulen, más inhibidor se libera para bloquear la división celular y la diferenciación en la caliptra. Además de ese inhibidor, las células del borde producen antibióticos para repeler patógenos, y/o señales que estimulan la simbiosis con bacterias u hongos. Junto con el mucílago y otros exudados, son la fuente de carbono que las plantas aporta a la rizosfera29 .
No se conoce la razón o ventaja evolutiva de por qué las células del borde necesitan desprenderse del resto de la raíz, sin embargo se ha planteado la hipótesis de que podrían ser buenas para mantener a los patógenos separados de la raíz, o permitir a hongos crecer cerca mientras se estabiliza un canal de comunicación entre estos29 .
Estructura secundaria

Los tejidos secundarios de la raíz son iguales a los tejidos secundarios del tallo, si bien difieren en el lugar en el que aparece el cámbium, ya que la organización de los tejidos vasculares primarios de ambos órganos es distinta. En la raíz, el cámbium se inicia a partir de células procambiales no diferenciadas y en forma de arcos sobre el borde interno del floema. Luego, a partir de las células más internas del periciclo, se forman nuevos arcos por fuera de los polos de xilema. Estos arcos se unen a los anteriores de modo que forman una capa continua que, en sección transversal, tiene forma sinuosa. Finalmente adquiere una forma cilíndrica, debido a que el xilema secundario se deposita más rápidamente sobre el lado interno del floema que en el exterior del protoxilema. En algunas especies, el cámbium que se origina en el periciclo forma radios medulares anchos. Los tejidos vasculares secundarios forman un cilindro continuo que incluye completamente al xilema primario. El floema primario es aplastado y algunas de sus células se diferencian en fibras.32
El xilema secundario de la raíz presenta menor cantidad de fibras, los vasos de tamaño uniforme, una escasa diferenciación de anillos de crecimiento, más elementos parenquimáticos vivos con función de reserva, más almidón y menos taninos que el xilema secundario del tallo. Generalmente en la raíz hay mayor cantidad de floema secundario en relación con la cantidad de leño que la que se observa en el tallo.32
Peridermis
La peridermis es el revestimiento del cuerpo vegetativo secundario de la planta que sustituye en su función a la epidermis y que se origina por la actividad de un meristema denominado felógeno. En la mayoría de las raíces el felógeno se produce en las células más externas del periciclo y entonces la peridermis es profunda lo que determina el desprendimiento del córtex cuando la raíz sufre crecimiento en grosor. El felógeno también puede originarse cerca de la superficie en algunos árboles y herbáceas perennes en las que el córtex cumple funciones de almacenamiento. En este caso la peridermis es superficial y el córtex se conserva.33 En Citrus la primera peridermis es subsuperficial y las siguientes son profundas. En las raíces de la palmera Phoenix dactylifera hay estructuras lenticelares que forman una especie de collar en torno a las raíces de menor orden.32Crecimiento secundario atípico
Varias raíces almacenadoras presentan crecimiento secundario atípico. En la zanahoria y en la remolacha azucarera (Beta vulgaris var. altissima) la raíz principal constituye la parte principal del órgano acumulador de reservas. En el primer caso, la masa principal del tejido reservante está formada por parénquima cortical. En la remolacha azucarera, en cambio, el engrosamiento se produce gracias a la formación sucesiva de varios anillos de cámbium en el parénquima cortical, las cuales se reconocen en el corte transversal debido al color más claro de los elementos leñosos a los que dan lugar. El enorme aumento de diámetro de la remolacha se debe a divisiones celulares y agrandamiento celular que ocurren simultáneamente en todos los anillos, gracias a la limitada formación de elementos xilemáticos lignificados y a la abundancia de células parenquimáticas. La sustancia de reserva, acumulada en el parénquima, es la sacarosa.34 35En las raíces de batata, Ipomoea batatas, el crecimiento secundario se inicia normalmente, luego se forman cámbiumes adicionales dentro del xilema secundario en torno a los vasos individuales o a grupos de vasos, a partir del parénquima paratraqueal. Estos cámbiumes producen algunos vasos hacia adentro y algunos elementos cribosos hacia afuera, pero sobre todo producen abundantes células parenquimáticas de almacenamiento. El proceso se repite indefinidamente; en estas raíces no se forma una corteza. Presentan laticíferos articulados simples. En el rabanito y otras brasicáceas el primer cámbium es normal, luego prolifera el tejido parenquimático del xilema y allí surgen cámbiumes anómalos que producen tejidos vasculares.35 En Dahlia la tuberización se produce por hipertrofia del xilema secundario, los vasos están dispersos en abundante parénquima leñoso que acumula inulina. Daucus carota almacena principalmente glucosa en las células del parénquima floemático secundario hipertrofiado.32
Modificaciones morfoanatómicas de la raíz
- Artículo principal: Morfología de la raíz

Raíces reservantes
Se presentan principalmente en plantas vivaces, es decir, aquellas donde los órganos aéreos desaparecen durante las épocas adversas, y también en muchas plantas bianuales. En éstas últimas, el órgano de reserva se forma durante el primer año, con una parte aérea muy corta, y en el segundo año el tallo se alarga y produce flores para lo que se utilizan las reservas almacenadas en la raíz. Desde el punto de vista anatómico, existen distintas variaciones estructurales en las raíces reservantes, pero en todas ellas se presenta abundancia de parénquima de reserva. Entre los tipos de raíces reservantes se encuentran:- Raíces napiformes, son aquellas raíces principales (axonomorfas) que se engrosan total o parcialmente por acumulación de sustancias de reserva. Consecuentemente, este tipo de raíces sólo se producen en dicotiledóneas. La zanahoria (Daucus carota) y el nabo (Brassica rapa) son ejemplos de este tipo de raíces.18 La mayoría de las veces interviene una gran parte del hipocótilo en su constitución por lo que estos órganos pueden resultar morfológicamente heterogéneos y, a pesar de su semejanza externa, pueden presentar considerables diferencias en su estructura anatómica. Así, en las raíces caulinotuberosas el engrosamiento ocurre tanto en la raíz principal como en el hipocótilo. La remolacha forrajera (Beta vulgaris var. crassa) y el rábano son ejemplos de este tipo de raíz reservante. En el caso del colinabo (Brassica napobrassica) y del apio (Apium graveolens) el segmento del tallo que sigue al hipocótilo, y que incluso presenta hojas normales, también se halla involucrado en el engrosamiento.18

- Raíces tuberosas: este término se utiliza cuando no solo la raíz principal, sino también las secundarias, o la porción apical de las raíces adventicias provenientes de los nudos inferiores del tallo, acumulan sustancias de reserva y agua, apareciendo todas engrosadas formando tubérculos radicales. Este tipo de raíz es característico de Dahlia, la yuca o mandioca (Manihot esculenta), Ranunculus ficaria y de algunas especies de marantáceas tropicales, como por ejemplo, el lairén (Calathea allouia). Este cultivo poco conocido presenta un rizoma y un sistema radical fibroso, con raíces duras y retorcidas, en el extremo de las cuales se forman las raíces tuberosas, elipsoidales a ovoides que constituyen la parte comestible de la planta. Estas raíces tuberosas miden de 1 a 5 cm de largo por 0,5 a 3 cm de ancho, están cubiertos por una cáscara dura, amarilla y brillante, con protuberancias espinosas. Debajo de la cáscara se encuentra el tejido parenquimatoso color claro a blanquecino que contiene almidón, encontrándose el centro generalmente vacío36 Los tubérculos radicales se asemejan a los tubérculos caulinares —como el de la papa— ya que, de hecho, son órganos análogos, pero su homología respecto a las raíces se reconoce porque poseen cofia, carecen de yemas o cicatrices foliares y por su estructura anatómica.18
- Xilopodios: este término se utiliza cuando la raíz principal de un sistema radical axonomorfo se lignifica y reserva agua. Este tipo de raíces es característico de algunas especies subarbustivas de sabanas, donde las precipitaciones son reducidas, razón por la cual se desarrolla tejido que reserva agua (parénquima acuífero) en la raíz. Ejemplo: añil (Indigofera suffruticosa).
Raíces especializadas como órganos de sostén y fijación
- Raíces contráctiles: son raíces adventicias, carnosas, largas y turgentes, las cuales se presentan en una cierta etapa del desarrollo de la planta y arrastran al brote cerca o debajo de la superficie del suelo, a un nivel adecuado para su desarrollo. En algunas plantas bulbosas, las raíces contráctiles están muy especializadas y provocan el desplazamiento del bulbo a mayor profundidad del suelo; estas raíces están poco lignificadas y presentan abundante parénquima, teniendo una duración de uno a tres años, al término de los cuales, las reservas se consumen y la raíz se acorta en un 30-40%.
- Raíces fúlcreas o zancos: son raíces que se originan en la base del tallo y se extienden en forma de arco hasta el suelo, actuando como soporte. Este tipo de raíces se presenta en el maíz, en algunas palmas y en los mangles (Rhizophora). El valor adaptativo de las raíces zancos no ha sido totalmente explicado, pero es evidente su importancia como soporte mecánico de árboles altos e inestables en suelos suaves y poco profundos. Asimismo, estas raíces pueden contribuir a la nutrición de algunas plantas cuyas raíces subterráneas han sido dañadas, en suelos inundados o pobremente aireados.
- Raíces respiratorias o neumorrizas: algunas plantas palustres presentan neumatóforos, raíces especializadas con geotropismo negativo. En el caso de Ludwigia peploides sobre los estolones se originan dos tipos de raíces: los neumatóforos y otras raíces adventicias, con geotropismo positivo. Los neumatóforos presentan la superficie rugosa, la estela es muy pequeña, y el córtex está formado por un aerénquima particular, constituido por células alargadas dispuestas en capas concéntricas.26

- Raíces haustoriales: son raíces muy modificadas que se presentan en plantas parásitas y hemiparásitas y están relacionadas con el tipo particular de nutrición de estas plantas. El origen de estas raíces puede ser embrional o adventicio. Las raíces haustoriales son el órgano de fijación y de absorción de estas plantas y representan los sitios en que se establece el contacto entre la planta huésped y la hospedera, considerándose: “órganos chupadores”.
Otras modificaciones en relación con el ambiente
Las plantas pueden presentar diversas adaptaciones, que responden a las distintas condiciones ambientales en que éstas se desarrollan. Una de las adaptaciones de las raíces a ambientes con escaso suministro de nutrientes en el suelo, especialmente el fósforo, son las raíces proteoides. Este tipo de raíces, denominadas también raíces proteiformes o raíces en racimo,37 son densos conglomerados de raíces laterales cortas y densamente espaciadas. Los conglomerados proteiformes se producen a intervalos de tiempo y distancias variables según la especie. Pueden formar matas de dos hasta cinco centímetros de espesor justo debajo de la superficie del suelo. Mejoran la absorción de nutrientes, posiblemente modificando químicamente el ambiente del suelo para mejorar la solubilización de los mismos. Como resultado, las plantas con raíces proteiformes puede crecer en suelos con escasa cantidad de nutrientes, como los suelos con deficiencia de fósforo, por lo que están presentes en plantas que son colonizadoras de ambientes xéricos.38Rizosfera

Las plantas secretan varios compuestos en la rizosfera que cumplen diversas funciones. Las estrigolactonas, por ejemplo, secretadas y detectadas por los hongos que participan de las micorrizas, estimulan la germinación de las esporas e inician cambios en las micorrizas que les permite colonizar las raíces. La planta parásita Striga también percibe la presencia de las estrigolactonas y sus semillas sólo germinan cuando este tipo de compuestos están presentes. Los rizobios también detectan la presencia de un compuesto secretado por las raíces hacia la rizosfera, el cual todavía no ha sido identificado con certeza, pero que desencadena los procesos que llevan a su colonización de los pelos radiculares y, por ende, a la formación de nódulos.
Algunas plantas secretan aleloquímicos desde sus raíces los que inhiben el crecimiento de otros organismos. Por ejemplo, la especie invasora Alliaria petiolata (brasicáceas) produce una sustancia que se cree que inhibe el desarrollo de asociaciones mutualísticas de otras especies en los bosques templados de Norteamérica.41
Micorrizas


Las ectomicorrizas son características de ciertos grupos de árboles y arbustos de regiones templadas, como lo son varios representantes de las fagáceas (la familia de los robles), las salicáceas (familia de los álamos y sauces), pináceas (pinos y cedros) y géneros como Eucalyptus y Nothofagus. El hongo crece entre las células de la raíz, rodeándolas sin penetrarlas, formando una estructura característica conocida como «red de Hartig». Además las raíces están rodeadas por una vaina formada por el hongo, llamada «manto fúngico»; las hormonas que secreta el hongo provocan la ramificación de la raíz, que adopta un aspecto esponjoso y ramificado. El micelio se extiende mucho hacia el suelo. Los pelos absorbentes a menudo están ausentes, siendo su función reemplazada por las hifas fúngicas.45 Los hongos que forman ectomicorrizas son en su mayoría basidiomicetes, pero hay también muchos ascomicetes. Las asociaciones son muy específicas, pero sin embargo Pinus sylvestris, por ejemplo, puede formar ectomicorrizas con 25 especies de hongos.18
Nódulos radicales


El nitrógeno molecular componente mayoritario de la atmósfera, es inerte y no aprovechable directamente por la mayoría de los seres vivos. La fijación de nitrógeno es el proceso por el cual este elemento se combina con oxígeno o con hidrógeno para dar óxidos o amonio que pueden incorporarse a la biosfera. Estas reacciones ocurren de forma abiótica en condiciones naturales como consecuencia de las descargas eléctricas o procesos de combustión y el agua de lluvia se encarga de arrastrar al suelo los compuestos formados, o bien se derivan de la síntesis química realizada en la industria de fertilizantes con un alto consumo de energía. La reducción de este elemento a amonio llevada a cabo por bacterias en vida libre o en simbiosis con algunas especies de plantas se conoce como fijación biológica del nitrógeno (FBN).
La fijación biológica del nitrógeno es un proceso muy antiguo que probablemente se originó en el Eón arqueano bajo las condiciones de ausencia de oxígeno de la atmósfera primitiva. Es exclusivo de Euryarchaeota y en seis de los más de 50 phyla de bacterias. Algunos de estos linajes coevolucionaron conjuntamente con las angiospermas estableciendo las bases moleculares de una relación de simbiosis mutualista. Los nódulos son las estructuras especializadas dentro de las cuales se lleva a cabo la FBN que algunas familias de angiospermas han desarrollado, principalmente en la corteza radicular. Las fanerógamas que han coevolucionado con diazótrofos actinorrícicos o con los rizobios para establecer su relación simbiótica pertenecen a once familias que se agrupan dentro del clado Rosidae de la filogenia molecular del gen rbcL que codifica parte de la enzima RuBisCO en el cloroplasto. Este agrupamiento indica que probablemente la predisposición a la formación de nódulos surgió una sola vez en las angiospermas y podría ser considerado como un carácter ancestral que se ha conservado o perdido en ciertos linajes. Sin embargo, la distribución tan dispersa de familias y géneros nodulantes dentro de este linaje, indica que la nodulación tiene múltiples orígenes.9
Los rizobios entran en los pelos radicales, que se deforman. La bacteria degrada la pared celular del pelo radicular y la penetra; el crecimiento del pelo se altera y se forma hacia adentro una estructura tubular llamada «hebra de infección». La hebra se dirige a la base del pelo y a través de las paredes celulares va al interior del córtex. Las bacterias inducen la división celular en las células corticales, las cuales se vuelven meristemáticas. Cuando los rizobios son liberados de las hebras de infección y penetran en las células radicales, quedan envueltos por invaginaciones de la membrana plasmática de los pelos radicales. Debido a la continua proliferación de bacteroides (rizobios desarrollados) y células corticales, se forman unos crecimientos tumorales que constituyen los nódulos.46
Cada nódulo presenta una región central con células invadidas por bacteroides de Rhizobium, y una zona meristemática que agrega nuevas células permitiendo al nódulo crecer y fijar nitrógeno indefinidamente. El nódulo está inervado por 1 a 5 trazas xilemáticas derivadas de la estela de la raíz. Cerca de la periferia del nódulo se forma una endodermis del nódulo, conectada con la endodermis de la raíz. Los nódulos están formados por células infectadas por Rhizobium que adquieren gran tamaño y por células más pequeñas, no infectadas, que son ricas en peroxisomas y retículo endoplasmático. Estas últimas tienen en la soja un papel muy importante en la producción de ureidos, compuestos derivados de la urea, a partir del nitrógeno molecular recién fijado por las células infectadas.47
En los alisos (Alnus) y otros árboles de las familias de las betuláceas, casuarináceas, eleagnáceas, miricáceas, ramnáceas, rosáceas y ulmáceas, las raíces se asocian simbióticamente con actinomicetes del género Frankia, que fijan nitrógeno. Forman nódulos en los cuales las bacterias llenan las células. Probablemente la nitrogenasa, enzima fijadora de nitrógeno, se localiza en la periferia de esas células.9 47
véase también Morfogénesis de los nódulos radiculares en leguminosas
Fisiología
Crecimiento radicular
El crecimiento radicular está determinado por factores genéticos y del medio. Entre los factores del medio se destacan las propiedades físicas y químicas del suelo, además de las variables que afectan el desarrollo aéreo del cultivo. Estas últimas pueden alterar drásticamente el crecimiento de las raíces debido a su dependencia de los productos de fotosíntesis proporcionados por la parte aérea.48 Desde un punto de vista físico, el crecimiento radicular depende en gran medida del agua. El potencial de presión, ,
o turgencia en el interior de las células, proporciona la fuerza
necesaria para la elongación celular. En el tejido en elongación, la
componente axial de esta fuerza es la que empuja la caliptra a través
del suelo.49
,
o turgencia en el interior de las células, proporciona la fuerza
necesaria para la elongación celular. En el tejido en elongación, la
componente axial de esta fuerza es la que empuja la caliptra a través
del suelo.49En el crecimiento radicular, el cual depende de la elongación celular, el agua tiene una participación fundamental al proporcionar la fuerza que produce la expansión de las paredes celulares. Esta acción del agua se puede describir conceptualmente mediante una ecuación originalmente por J.A. Lockhart50 y utilizada ampliamente con posterioridad en estudios de elongación celular.51 52 53 La ecuación establece que la tasa de elongación celular es proporcional al potencial de presión que se desarrolla en el interior de la célula en elongación por encima de un determinado valor umbral. Ese valor umbral es el mínimo potencial de presión (
) que se requiere para producir una deformación plástica en las paredes celulares. La ecuación es de la siguiente forma:
 es el potencial de presión mínimo necesario para que se produzca una deformación plástica a nivel celular.54
es el potencial de presión mínimo necesario para que se produzca una deformación plástica a nivel celular.54Tanto Eg como
son variables.55
Para el caso de crecimiento radicular, la ecuación anterior debe
modificarse para incluir el impedimento mecánico del suelo al proceso de
crecimiento. El impedimento mecánico puede visualizarse como una
presión desde el suelo hacia la raíz, es decir, como una reacción del
suelo ante su deformación impuesta por el crecimiento de la raíz, la que
puede describirse matemáticamente como:52![TE = E_{g} [{\Psi_{p} - (\Psi_{p,u} + \Sigma_{n}})]](http://upload.wikimedia.org/math/2/6/0/260ae0f73f93849b2dbaf2af4afa1945.png)
 es la presión externa del suelo sobre las células en crecimiento de la raíz. El valor de no se puede medir directamente y para células en elongación se supone que vale
es la presión externa del suelo sobre las células en crecimiento de la raíz. El valor de no se puede medir directamente y para células en elongación se supone que vale  donde
donde  es el potencial total del agua y
es el potencial total del agua y  ,
el potencial de solutos. El potencial total del agua es el potencial
químico del agua en un sistema o parte de un sistema, expresado en
unidades de presión y comparado con el potencial químico del agua pura, a
la presión atmosférica y a la misma temperatura y altura, con el
potencial químico del agua tomado como referencia e igual a cero. El
potencial osmótico o de solutos es la fracción del potencial total que
resulta del efecto de la interacción de las moléculas de agua con todos
los solutos presentes en ella. Su valor es igual a cero para el agua
pura, y negativo e inversamente proporcional a la concentración de sales
disueltas, es decir, a mayor cantidad de sales, el valor aumenta
negativamente, lo que significa que la presencia de iones en el agua
reduce la energía de ésta.56 Reemplazando el valor de en la ecuación anterior se obtiene:
,
el potencial de solutos. El potencial total del agua es el potencial
químico del agua en un sistema o parte de un sistema, expresado en
unidades de presión y comparado con el potencial químico del agua pura, a
la presión atmosférica y a la misma temperatura y altura, con el
potencial químico del agua tomado como referencia e igual a cero. El
potencial osmótico o de solutos es la fracción del potencial total que
resulta del efecto de la interacción de las moléculas de agua con todos
los solutos presentes en ella. Su valor es igual a cero para el agua
pura, y negativo e inversamente proporcional a la concentración de sales
disueltas, es decir, a mayor cantidad de sales, el valor aumenta
negativamente, lo que significa que la presencia de iones en el agua
reduce la energía de ésta.56 Reemplazando el valor de en la ecuación anterior se obtiene: y
son valores negativos, en consecuencia, de acuerdo a la ecuación
anterior la elongación celular en las raíces se producirá sólo si se
cumple la relación < (
y
son valores negativos, en consecuencia, de acuerdo a la ecuación
anterior la elongación celular en las raíces se producirá sólo si se
cumple la relación < ( )
)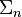 siempre y cuando se cumpla la condición descripta en esa inecuación.
Este comportamiento de los componentes del potencial del agua se ha
observado en diferentes especies: en raíces de plántulas de arveja52 y en raíces de maíz.54
Cuando ocurren deficiencias hídricas, el potencial agua de la raíz
disminuye pero también disminuye ψs debido a un aumento del nivel de los
solutos en las células.54
En síntesis, la exploración del suelo por las raíces necesita de una
turgencia en las células en expansión. Esa turgencia debe proporcionar
una fuerza tal que produzca deformaciones plásticas en las células y
venza el impedimento mecánico que opone el suelo. Aparentemente, un
proceso de ajuste osmótico de las raíces (disminución del valor de ψs)
sería un mecanismo que permitiría el crecimiento radicular aun a
potenciales de agua bajos en el suelo, es decir, cuando ocurren
deficiencias hídricas.54
siempre y cuando se cumpla la condición descripta en esa inecuación.
Este comportamiento de los componentes del potencial del agua se ha
observado en diferentes especies: en raíces de plántulas de arveja52 y en raíces de maíz.54
Cuando ocurren deficiencias hídricas, el potencial agua de la raíz
disminuye pero también disminuye ψs debido a un aumento del nivel de los
solutos en las células.54
En síntesis, la exploración del suelo por las raíces necesita de una
turgencia en las células en expansión. Esa turgencia debe proporcionar
una fuerza tal que produzca deformaciones plásticas en las células y
venza el impedimento mecánico que opone el suelo. Aparentemente, un
proceso de ajuste osmótico de las raíces (disminución del valor de ψs)
sería un mecanismo que permitiría el crecimiento radicular aun a
potenciales de agua bajos en el suelo, es decir, cuando ocurren
deficiencias hídricas.54Desarrollo
Una característica básica de las raíces es su patrón radial de desarrollo, es decir, que está formada por anillos concéntricos (o capas) de diferentes tejidos. Los tres tipos fundamentales son la epidermis, el parénquima cortical y el tejido vascular. La raíz primaria del organismo modelo Arabidopsis thaliana presenta anillos de una sola célula de espesor de epidermis, cortex, endodermis y periciclo rodeando el cilindro vascular.57 Como en muchas otras especies, cada una de las filas de células de la raíz de Arabidopsis se origina en un conjunto específico de células en la región meristemática denominadas células iniciales las cuales, a su vez, rodean un grupo de células especializadas denominadas centro quiescente. Las iniciales conjuntamente con el centro quiescente conforman el promeristema de la raíz.58 El promeristema está compuesto por células que derivan, a su vez, de dos células -una apical y otra basal- que se generan durante la primera división del cigoto durante la embriogénesis. La célula basal da origen al centro quiescente y a las regiones de la columela y de la caliptra, mientras que la célula apical origina las restantes células del promeristema. De hecho, la arquitectura del promeristema de la raíz queda definida durante el estado de corazón durante la embriogénesis.59 Mediante experimentos que hacen uso de manipulaciones físicas —ablación por láser de células específicas— y genéticas —análisis de mutantes que exhiben diferentes patrones anormales de desarrollo—60 se ha demostrado que la identidad de cada tejido dentro de la raíz está determinada por la ubicación de las células más que por su origen.61 En otras palabras, las células de la raíz muestran un gran plasticidad de desarrollo. De hecho, cada célula que comienza a diferenciarse para dar origen a uno de los tres tejidos fundamentales (epidermis, parénquima o vascular) puede cambiar su programa de desarrollo debido a modificaciones en su posición relativa o a factores ambientales externos.62 63Absorción de agua

El sistema de raíces pueden absorber agua del suelo siempre y cuando el potencial hídrico de las raíces sea más bajo que el potencial hídrico del suelo circundante. La absorción de agua
 durante una unidad de tiempo es proporcional a la diferencia de potencial hídrico entre la raíz (
durante una unidad de tiempo es proporcional a la diferencia de potencial hídrico entre la raíz ( ) y el suelo (
) y el suelo ( ) e inversamente proporcional a la resistencia (
) e inversamente proporcional a la resistencia ( ) que opone el suelo a la transferencia de agua, como se describe en la siguiente ecuación:64
) que opone el suelo a la transferencia de agua, como se describe en la siguiente ecuación:64
La ósmosis se realiza siempre desde una región con potencial hídrico alto (valores negativos pequeños de
) a otra región de bajo potencial hídrico (valores negativos grandes de ). Por ejemplo, la ósmosis ocurrirá desde el suelo con un valor = - 3 MPa a la raíz con un valor =
- 4 MPa. En la mayoría de los casos, la solución del suelo es muy
diluida, de manera que el potencial osmótico suele ser pequeño. Sin
embargo, algunos suelos de regiones áridas contienen elevadas
concentraciones de sales solubles y reciben el nombre de suelos salinos.
La absorción de agua por las raíces en esos casos supone la adquisición
por parte de las plantas de mecanismos fisiológicos particulares que
les permiten adaptarse a tales condiciones. Las raíces desarrollan
usualmente un potencial desde unas pocas décimas hasta -1 MPa, mientras
que las plantas adaptadas a ambientes áridos (xerófitas) pueden desarrollar hasta -6 MPa56El movimiento del agua y los minerales del suelo se inicia primero a través de los pelos absorbentes y luego pasa al cortex, que es una capa de células de parénquima que tiene espacios intercelulares. Hay tres vías disponibles para el movimiento del agua en el cortex. A través de las paredes celulares, a través del citoplasma de las células vía plasmodesmos y a través del citoplasma y vacuolas de las células del cortex. El flujo a través de las vías señaladas es inversamente proporcional a la resistencia que encuentre el agua. El camino más utilizado es a través de las paredes celulares (apoplasto). En tanto que a través del citoplasma (simplasto), la resistenda al agua es mayor debido a la alta viscosidad relativa del citoplasma; sin embargo, la ciclosis parece ayudar al movimiento del agua. La tercera vía que involucra el movimiento a través de capas de membranas del citoplasma y las vacuolas, presenta mayor resistencia que la segunda vía, por lo que esta ruta no es muy utilizada.
Cualquiera que sea el camino empleado, el agua llega hasta la endodermis, donde el movimiento es restringido por la banda de Caspari. El agua y las sales minerales deben cruzar esta barrera a través del protoplasto de una célula endodérmica, cuya membrana plasmática diferencialmente permeable, retarda la difusión de los iones. Debido a esta banda, las sustancias que se mueven por la vía aploplástica continúan su camino por el simplasto, en el que ingresan siguiendo un gradiente de potencial electroquímico. Una vez que se cruza esta barrera, el agua ingresa a la estela, donde de nuevo tiene resistencias similares a las observadas en el cortex, allí otra vez el agua se moverá preferiblemente a través de las paredes celulares basta entrar a los vasos del xilema y traqueidas, los principales tejidos conductores de agua en las plantas superiores. A nivel de la membrana celular, el agua y los minerales ingresan en la célula a través de dos sistemas de transporte: los canales y los transportadores. Los transportadores constituyen una familia compleja de proteínas; algunos están involucrados en la difusión facilitada, otros participan en el transporte activo. Una vez que los iones atraviesan la endodermis, pasan al xilema en forma pasiva o activa y, junto con el agua, la corriente transpiratoria los transporta hacia la parte aérea de la planta. Durante el camino, los iones son cedidos al tallo y a las hojas.65
Movimientos de la raíz
Geotropismo
La gravedad es un estímulo ambiental único, en el sentido de que está presente de manera continua, es unidireccional y tiene una intensidad constante. Las plantas han desarrollado mecanismos que utilizan este indicador ambiental para guiar su dirección de crecimiento y tener acceso óptimo a recursos esenciales. Este tropismo asegura que las raíces crezcan hacia abajo en el suelo (gravitropismo positivo), donde pueden tomar agua y minerales, mientras que los vástagos crecen hacia arriba, donde pueden fotosintetizar, reproducirse y dispersar las semillas (gravitropismo negativo). Para responder a este tropismo, la planta tiene que percibir su orientación en el campo gravitatorio. La percepción de un cambio en la orientación del órgano (graviestimulación) por las células perceptoras de la gravedad (estatocitos) da como resultado la formación de una señal bioquímica que se transmite al sitio del órgano donde se desarrolla la curvatura.66Charles Darwin en 1880 observó que cuando las raíces son bloqueadas por un obstáculo las puntas se aplanaban, adquirían una forma oblicua y se curvaban casi en ángulo recto para tomar una nueva dirección de crecimiento. Darwin postuló que durante la graviestimulación había «algo» que se movía desde la punta de la raíz a la zona en la que esta se curvaba.67 En la primera parte del siglo XX, N. Cholodny68 y F. Went69 propusieron independientemente que debido a la redistribución de una sustancia promotora del crecimiento de un lado de la raíz se producía la respuesta gravitrópica. Llamaron a esta sustancia auxina que, en griego, significa «aumentar», debido a sus propiedades promotoras de la elongación celular. La auxina se purificó muchos años después y llegó a ser la primera sustancia de las plantas que fue categorizada como una fitohormona. La señalización disparada por la gravedad, entonces, controla aspectos específicos del transporte de auxinas y, de hecho, mutaciones en los transportadores de auxina afectan al gravitropismo en tallos y en raíces.70
Según la hipótesis de los estatolitos-almidón, la percepción de la gravedad en las plantas está mediada por la sedimentación o la presión/tensión ejercida por los plástidos llenos de almidón (denominados estatolitos) dentro de las células de la columela de la cofia en la raíz y dentro de la capa de almidón de la endodermis en el tallo. Esta hipótesis se encuentra avalada por la observación de que la disminución —ya sea por causas genéticas o fisiológicas— del almidón en ambos tipos de células determina una alteración en la respuesta gravitrópica. Se desconoce el modo por el cual los estatolitos de la raíz o del tallo perciben la sedimentación de los amiloplastos. No obstante, una vez que el estímulo se percibe, se origina una señal asimétrica en las células de la columela, que produce un movimiento lateral de auxinas. La diferencia de concentración de auxinas en ambos lados de la raíz determina que la misma se curve hacia el lado de menor concentración debido a que, del otro lado, las células se elongan más.71 72
Hidrotropismo
La supervivencia de las plantas terrestres depende de la capacidad de las raíces para obtener agua y nutrientes del suelo. El crecimiento direccional de las raíces con relación a la disponibilidad de agua se conoce como hidrotropismo, y comienza en la cofia con la percepción del gradiente de humedad. Todavía no se sabe cabalmente el modo ni el tipo de célula de la cofia que percibe el gradiente de humedad. No obstante, el uso de una mutante de Pisum sativum denominada ageotropum permitió establecer la independencia de las vías de percepción y señalización para ambos tipos de tropismos ya que sus raíces no responden a la gravedad pero si responden al hidrotropismo.73La capacidad de la cofia de percibir gradientes de humedad parece generar una señal dominante que debilita la percepción y respuesta al estímulo de la gravedad. Se ha observado que la respuesta reducida a la gravedad en raíces hidroestimuladas de Arabidopsis y rábano se debe en parte a la degradación simultánea de los amiloplastos en sus estatocitos. Las raíces estimuladas por tacto retardan la tasa de sedimentación de los amiloplastos, reduciendo también la respuesta gravitrópica. Sin embargo, la respuesta hidrotrópica ejerce un efecto más dramático que el tigmotropismo, quizás por la importancia del agua para la supervivencia de las plantas.74
Estos mecanismos indican que los estatocitos parecen haber adquirido durante su evolución diferentes tipos de receptores para percibir una gran variedad de estímulos que además son capaces de integrar cuantitativamente las diferentes señales e iniciar la respuesta trópica adecuada. Los estatocitos, consecuentemente, podrían utilizar elementos comunes de señalización, tales como redistribución de auxinas, cambios en el pH, concentración de Ca2+, en todos los sistemas perceptores y de ese modo sincronizar las diferentes respuestas.75
Evolución

A pesar que existen trazas de impresiones similares a raíces en suelos fosilizados del Silúrico tardío,77 los fósiles muestran que las primeras plantas carecían de raíces. Muchos presentaban zarcillos que se extendía a lo largo o debajo de la tierra, con ejes verticales o talos, y algunos incluso muestran ramas subterráneas no fotosintéticas que carecen de estomas. No obstante, la distinción entre una raíz y una rama especializada se basa en el desarrollo de las mismas, ya que las raíces verdaderas siguen una trayectoria de desarrollo diferentes a la de los tallos. Además, las raíces difieren en su patrón de ramificación, y en la posesión de un caliptra. Todos estos aspectos, lamentablemente, no aparecen en el registro fósil.10 Así, mientras que las plantas del Silúrico-Devónico —tales como Rhynia y Horneophyton— poseían el equivalente fisiológico de las raíces, las raíces propiamente dichas —definidas como órganos diferenciados de los tallos— no hicieron su aparición hasta períodos geológicos posteriores.10 Desafortunadamente, rara vez se conservan las raíces en el registro fósil por lo que la comprensión de su origen evolutivo es escasa.10 Los rizoides -pequeñas estructuras que realizan la misma función que las raíces, que por lo general presentan solo una célula de diámetro- probablemente evolucionaron más tempranamente, incluso antes de que las plantas colonizaran la tierra. De hecho, es un órgano que presentan las Characeae, un grupo de algas hermano a las plantas terrestres.10 Los rizoides probablemente evolucionaron más de una vez, los rizoides de los líquenes, por ejemplo, realizan una función similar.10

Sin embargo, ninguno de estos fósiles muestran todos los aspectos característicos de las raíces modernas.10 Las raíces y las estructuras similares a raíces se hicieron cada vez más comunes y de mayor longitud durante el Devónico, con árboles de Lycopodium que emitían raíces de alrededor de 20 cm de longitud en los períodos Eifeliano y Givetiano. Durante la etapa subsiguiente, el período Frasniense, los ejemplos mencionados fueron acompañados por las progimnospermas, que enraizaban hasta aproximadamente un metro de profundidad.13 Las verdaderas gimnospermas y algunos helechos también formaron sistemas de raíces poco profundas durante el período Fameniano tardío.13
Los denominados «rizomorfos» de los licopodios proporcionan un enfoque ligeramente diferente al de las plantas actuales con respecto al enraizamiento. Son equivalentes a tallos, con órganos equivalentes a hojas que desempeñan el papel de raicillas. Una arquitectura similar se observa en el género extinto Isoetes y Lepidodendron, por lo que parece que las raíces han evolucionado al menos en dos oportunidades diferentes e independientes: por un lado en las licófitas y, por el otro, en las demás plantas.10
Para el Devónico intermedio a tardío, la mayoría de los grupos de plantas desarrollaron un sistema radicular de alguna naturaleza.11 A medida que las raíces se hacían más largas, podían sustentar estructuras aéreas más altas y podían explorar el sustrato a mayor profundidad.12 Esta exploración más eficiente del suelo tuvo profundos efectos ecológicos: no solo permitió a las plantas la conquista de nuevos hábitats sino también la posterior colonización de los mismos por los animales y los hongos.13
Importancia de las raíces en la adaptación biológica y la evolución
Las raíces son un ejemplo de cómo los organismos pueden modificar el ambiente en el que se desarrollan, y en este caso particular, lo modificaron a escala global (con la alteración de los ciclos biogeoquímicos) y a nivel local (a través de las interacciones bióticas de la raíz). La evolución de los sistemas radiculares para anclaje, absorción de agua y adquisición de nutrientes, fueron esenciales para la diversificación de las plantas en tierra firme79 . No obstante, las raíces también tuvieron un impacto en las condiciones atmosféricas y la composición del sustrato. A través de la acción fisiológica de las raíces y la secreción de ácidos en la rizosfera, las plantas aceleraron la erosión de silicatos, el rompimiento de rocas y la introducción de enormes cantidades de material orgánico que llevaron al desarrollo del suelo que conocemos actualmente. Eso incrementó la tasa de reacción del CO2 atmosférico causando un cambio en el equilibrio del ciclo del carbono y reduciendo drásticamente el CO2 atmosférico, lo que finalmente llevó a un enfriamiento climático global79 .Por el efecto que tiene la rizosfera en el desarrollo de la raíz y por el efecto de esta en la dinámica de la rizosfera, las raíces tienen el potencial de influir directamente en la adaptación biológica de los organismos con los que interactúan, y por medio de ellos, también puede dirigir su propia evolución. El resultado de estas interacciones puede ser muy diverso, por ejemplo la interacción planta-patógeno puede llegar a ser más severa cuando se cultivan recurrentemente plantas susceptibles a patógenos ya que eso favorece la acumulación de estos últimos en el sustrato y por lo tanto ocasiona un incremento en su incidencia. En hábitats naturales, los patogenos de las raíces constituyen una fracción pequeña de la población total de organismos del sustrato y los brotes de plagas son raras. Es por eso que la práctica inadecuada de la agricultura amplía las interacciones perjudiciales de las raíces con los microorganismos. Sin embargo, también hay interacciones que favorecen la adaptación de ciertas especies al medio pero evitan la invasión de otras. Ese es el caso de la simbiosis hongo-raíz. En general se pueden distinguir ectomicorrizas y endomicorrizas como dos tipos de simbiosis hongo-raíz. Las plantas con endomicorrizas pueden ser de baja especificidad y no dependen totalmente del hongo para la ingesta de nutrientes pero su eficiencia para adquirir los recursos es baja. Donde hay árboles con ectomicorrizas, las endomicorrizas quedan fuera, ya que los primeros son más eficientes en la utilización de los recursos (por ejemplo en los desiertos). Finalmente, de acuerdo a la teória de juegos se ha planteado que el desarrollo los sistemas radiculares podría depender enormemente de la presencia de competidores, ya que las plantas con rasgos óptimos no serían evolutivamente estables a nivel de comunidad. Los modelos de teorías de juegos demuestran que la producción de mejores sistemas radiculares u hojas de mayor tamaño depende enormemente del nivel de reconocimiento de los vecinos, es decir si las plantas interactúan predominantemente con ellas mismas se esperaría menor desarrollo de estructuras óptimas80 . Una forma a través de la cual se puede dar este reconocimiento es por medio de interacciones entre raíces, ya sea física o a través de exudados o micorrizas, de esta manera las interacciones entre raíces puede favorece la adaptación de comunidades.
Las raíces de las plantas además de estar expuesta a los microorganismos que habitan el sustrato también son su fuente primaria de nutrientes. Es por eso que no sorprende que raíces y microorganismos hayan evolucionado métodos de interacción para inhibir patógenos o para atraer simbiontes a la rizosfera. Ya sea con la producción de células del borde, con la organización del meristemo apical de la raíz o con la secreción de exudados en la rizosfera, las raíces tienen el potencial de establecer diferentes relaciones bióticas por medio de las cuales modifican su entorno a diferentes escalas. A su vez, esas interacciones influyen en la plasticidad fenotípica de las plantas y en la adaptación de las especies con las que se relacionan. Gracias a sus raíces y a la habilidad de estas de modificar su nicho81 , las plantas pueden dirigir su propia evolución y afectar la de otras especies.
Importancia económica
Véase también: Usos medicinales de las raíces.





El empleo de las raíces en medicina humana y veterinaria es usual en muchas culturas alrededor de todo el mundo. Las raíces del ñame, por ejemplo, son una fuente de estrógenos utilizados en las píldoras para el control de la natalidad y desde las raíces de Rauwolfia serpentina se extrae el alcaloide reserpina. Las raíces del ginseng, del acónito y de la ipecacuana son también muy apreciadas por sus propiedades farmacológicas. El insecticida rotenona, que se extrae de las raíces de Lonchocarpus, es un ejemplo de otras de las utilidades que presentan las raíces.82 Las propiedades farmacológicas de las raíces de muchas especies están basadas en la producción y acumulación de metabolitos secundarios. Los extractos de plantas son el mayor recurso para la producción de productos farmacéuticos. Las técnicas de cultivo in vitro de células u órganos han permitido estandarizar la producción de metabolitos secundarios. Por otro lado, la ingeniería genética permite también incrementar la producción de tales sustancias. Así, la transformación genética mediada por Agrobacterium rhizogenes es un método de transformación que presenta fundamentalmente dos aplicaciones: la producción de raíces con alta tasa de crecimiento capaces de sintetizar metabolitos secundarios como productos farmacéuticos, aditivos alimentarios y cosméticos y, por otra parte, representa una valiosa herramienta para la estimulación de la formación de raíces en especies recalcitrantes, por ejemplo, en plantas leñosas. Este método de obtención de raíces genéticamente modificadas presenta muchas ventajas en comparación con el cultivo de células y de raíces sin transformar.83 Algunas de las cuales son un crecimiento más acelerado y una mayor productividad de metabolitos secundarios.84 Entre los metabolitos secundarios producidos por raíces genéticamente transformadas se encuentran los terpenoides,85 la hiosciamina y la escopolamina86
Las raíces de otras especies, finalmente, se emplean como material para realizar obras de arte o implementos para el hogar. Las raíces del ciprés calvo se venden como recuerdos, como grabados o como bases para lámparas en el arte popular. Los nativos americanos utilizan las raíces flexible del abeto blanco para cestería.82
Las raíces de muchas especies protegen el medio ambiente debido a que sostienen al suelo o la arena donde crecen, evitando de ese modo que el agua o el viento lo erosionen. Las raíces de los árboles, por otra parte, pueden levantar y destruir las aceras de hormigón y triturar u obstruir las tuberías enterradas.82 A continuación se presenta un listado de las especies cultivadas cuyas raíces se utilizan como alimento.
| Nombre científico | Nombre común | Familia |
|---|---|---|
| Justicia insularis | - | Acanthaceae |
| Apium graveolens var. rapaceum | apio nabo, apirrábano | Apiaceae (Umbeliferae) |
| Arracacia xanthorrhiza | arracacha | Apiaceae |
| Chaerophyllum bulbosum | perifollo tuberoso, perifollo bulboso | Apiaceae |
| Daucus carota | zanahoria | Apiaceae |
| Glehnia littoralis | - | Apiaceae |
| Myrrhis odorata | - | Apiaceae |
| Pastinaca sativa | pastinaca, chirivía | Apiaceae |
| Petroselinum crispum var. tuberosum | perejil tuberoso | Apiaceae |
| Sium sisarum | escaravia, chirivía de azúcar, escaravia tudesca, sisaro | Apiaceae |
| Arctium lappa | bardana mayor, lampazo | Asteraceae (Compositae) |
| Cichorium intybus | achicoria silvestre, achicoria de raíz, achicoria de bruselas, amargón, almirón | Asteraceae |
| Cirsium dipsacolepis | cardo comestible, cardo de huerta | Asteraceae |
| Polymnia sonchifolia | arboloco, jacón, llamón, aricoma, jiquimilla | Asteraceae |
| Scolymus hispanicus | escolimo, cardillo, almirón de España | Asteraceae |
| Scorzonera hispanica | escorzonera, escorzonera de España, salsifí negro | Asteraceae |
| Taraxacum officinale | diente de léon, taraxacón | Asteraceae |
| Tragopogon porrifolius | salsifí blanco | Asteraceae |
| Tragopogon pratensis | barba cabruna | Asteraceae |
| Armoracia rusticana | rábano rusticano, rábano picante, rábano silvestre | Brassicaceae (Cruciferae) |
| Brassica juncea var. megarrhiza | mostaza tuberosa | Brassicaceae |
| Brassica napus var. napobrassica | rutabaga, nabo sueco, nabo de Suecia | Brassicaceae |
| Brassica rapa var. rapa | nabo | Brassicaceae |
| Lepidium meyenii | maca | Brassicaceae |
| Raphanus sativus | rábano, rabanito, rabanete, rabanillo | Brassicaceae |
| Campanula rapunculus | rapónchigo, rapúnculo, ruiponce, raipóntico | Campanulaceae |
| Beta vulgaris | remolacha hortelana, remolacha roja, remolacha de mesa | Chenopodiaceae |
| Convolvulus japonicus | boniato del Japón | Convolvulaceae |
| Ipomoea batatas | batata, boniato, camote, papa dulce, moniato | Convolvulaceae |
| Manihot esculenta | yuca, tapioca, mandioca, guacamote | Euphorbiaceae |
| Bauhinia esculenta | bauhinia, judía de Marama | Fabaceae (Leguminosae) |
| Pachyrhizus ahipa | ahipa | Fabaceae |
| Pachyrhizus tuberosus | frijol alado, dólico de Goa | Fabaceae |
| Psophocarpus tetragonolobus | bauhinia, judía de Marama | Fabaceae |
| Pueraria lobata | kudzú común, kudzú tropical, hierba kudzú | Fabaceae |
| Calathea allouia | dale dale, agua bendita, cocurito, leren, sewi, lairén, yairen | Marantaceae |
| Euryale ferox | - | Nymphaeaceae |
No hay comentarios:
Publicar un comentario